O‘simliklar fiziologiyasi



![Katta markaziy vakuol , hujayra shirasi bilan to'ldirilgan va membrana
bilan chegaralangan bo'shliq - tonoplast [1] [2] . Vakuola hujayra
turgorini saqlashda muhim rol o'ynaydi , molekulalarning sitozoldan
hujayra sekretsiyasigacha harakatini nazorat qiladi, ozuqa
moddalarini saqlaydi va eski oqsillar va organellalarni parchalaydi .
Hujayra devori asosan tsellyulozadan , shuningdek , gemitsellyuloza ,
pektin va ko'p hollarda lignindan iborat . Hujayra membranasi
tepasida joylashgan protoplast tomonidan hosil bo'ladi . U xitindan
iborat zamburug'larning hujayra devoridan va peptidoglikan (murein)
dan qurilgan bakteriyalardan farq qiladi.
Hujayralar o'rtasidagi aloqaning ixtisoslashgan yo'llari -
plazmodesmata [3] , sitoplazmatik ko'priklar: qo'shni hujayralarning
sitoplazmasi va endoplazmatik retikulum (ER) hujayra devoridagi
teshiklar orqali aloqa qiladi](/data/documents/3285ee6e-ff79-4fda-ab05-690dee8b5594/page_5.png)



















O ‘ simliklar fiziologiyasi
O ‘ simliklar fiziologiyasining asosiy yo ‘ nalishlari O simliklar fiziologiyasi — o simliklar hayot faoliyati umumiy ʻ ʻ qonuniyatlarini o rganadigan fan. O simliklar fiziologiyasi ʻ ʻ o simliklarning o z tanasini tiklashi, ko payishi uchun zarur ʻ ʻ ʻ bo lgan mineral moddalar va suvni o zlashtirishi, o sishi, ʻ ʻ ʻ rivojlanishi, gullashi va meva hosil qilishi, ildizdan (mineral) va havodan (fotosintez) oziqlanishi, nafas olishi, biosintez qilishi, zaxira moddalarni to plashi va boshqalar ʻ jarayonlarni o rganadi. O simliklar fiziologiyasi hayotiy ʻ ʻ jarayonlarning tashqi muhit bilan bog liqligini ochib berish ʻ orqali o simliklarning umumiy mahsuldorligi, oziq qiymati, ʻ organ va to qimalarining texnologik sifatini oshirish usuli va ʻ metodlarining nazariy asoslarini tadqiq qiladi. Fiziologik tadqiqotlar o simliklarning o sishi va mo l hosil berishi uchun ʻ ʻ ʻ zarur bo lgan tuproq va iqlim sharoitlari hamda ulardan ʻ oqilona foydalanishning nazariy asosi hisoblanadi.
O simliklar fiziologiyasiga oid masalalar uning ob yekti bo lgan ʻ ʼ ʻ yashil o simliklarning o ziga xos xususiyatlari bilan aniqlanadi. ʻ ʻ Yashil o simliklar quyosh nuridan energiya manbai sifatida ʻ foydalanishi va uni organik birikmalarning kimyoviy energiyasiga aylantirishi, ya ni fotosintez jarayonini amalga ʼ oshirishi bilan boshqa organizmlardan farq qiladi. O simliklarning o ziga xos xususiyatlari ularning umumiy ʻ ʻ anatomik va morfologik tuzilishi bilan chambarchas bog liq. ʻ O simliklar organizmi, odatda, yer ustki va yer ostki ʻ qismlarining shoxlanishi tufayli juda katta sathni egallaydi. Bu holat ularning tuproq va havoning katta hajmi bilan bog lanishiga imkon beradi. O simliklarda doimiy, o zgarmas ʻ ʻ ʻ ichki muhit bo lmasligi tufayli ular to qimalarining harorati, ʻ ʻ kislorod va karbonat angidrid miqdori hamda boshqa ko rsatkichlari o zgarib turadi. ʻ ʻ
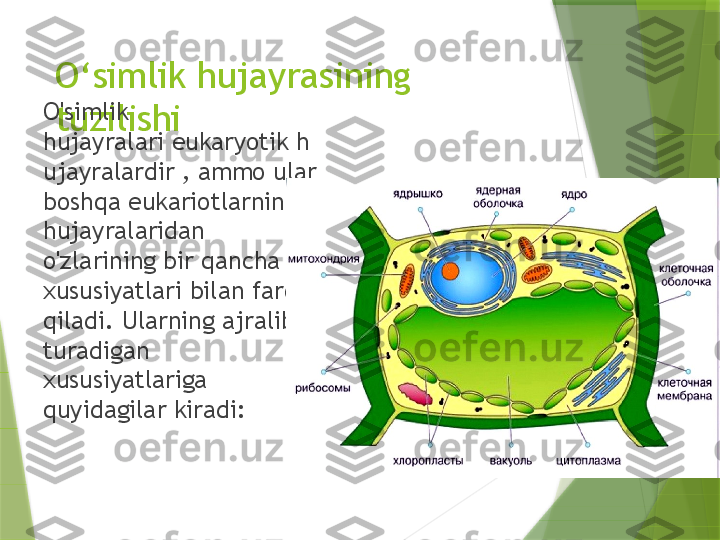
O ‘ simlik hujayrasining tuzilishiO'simlik hujayralari eukaryotik h ujayralardir , ammo ular boshqa eukariotlarning hujayralaridan o'zlarining bir qancha xususiyatlari bilan farq qiladi. Ularning ajralib turadigan xususiyatlariga quyidagilar kiradi:
Katta markaziy vakuol , hujayra shirasi bilan to'ldirilgan va membrana bilan chegaralangan bo'shliq - tonoplast [1] [2] . Vakuola hujayra turgorini saqlashda muhim rol o'ynaydi , molekulalarning sitozoldan hujayra sekretsiyasigacha harakatini nazorat qiladi, ozuqa moddalarini saqlaydi va eski oqsillar va organellalarni parchalaydi . Hujayra devori asosan tsellyulozadan , shuningdek , gemitsellyuloza , pektin va ko'p hollarda lignindan iborat . Hujayra membranasi tepasida joylashgan protoplast tomonidan hosil bo'ladi . U xitindan iborat zamburug'larning hujayra devoridan va peptidoglikan (murein) dan qurilgan bakteriyalardan farq qiladi. Hujayralar o'rtasidagi aloqaning ixtisoslashgan yo'llari - plazmodesmata [3] , sitoplazmatik ko'priklar: qo'shni hujayralarning sitoplazmasi va endoplazmatik retikulum (ER) hujayra devoridagi teshiklar orqali aloqa qiladi