SENSOR TIZIMLAR VA ULARNING FUNKSIYASI









































SENSOR TIZIMLAR VA ULARNING FUNKSIYASI Reja: 1. Sensor tizimlarning morfo-funksional tuzilishi va xususiyatlari; 2. Retseptorlarning tasnifi; 3. Sensor tizimlarning xususiy morfofunksional tuzilishi; 4. Ko‘rish vestibulyar va harakat analizatorlarining fiziologiyasi.
.1. Sensor tizimlar, ularningfunksiyalari, umumiy tuzilishi va xususiyatlari Sensor tizimlar tarkibida bir necha qavat neyronlar mavjud. SHu sababli sensor tizimlarning asosiy shakllanish prinsipi bu ko‘p qavatlikdir. Boshlang’ich neyronlar reseptorga qarashli, oxirgi neyronlar esa miya po’stlog’ining assosiativ mintaqasidagi neyronlarga ko‘shiladi (5, 7, 4, 6, 1, 2 maydonlar). Ko’p qavatlilik bosh miya po’stlog’ining polimodal (turli xarakterga ega bo’lgan) tabiatini ifodalaydi, chunki polimodal signallarning birlamchi afferent sentezi markazi assosiativ mintaqalar hisolanadi (Ayrapetyans, Batuyev). Ko’p kanallik – organizmning tashqi va ichki muhitidan turli kanallar orqali bir-biriga o’xshamagan turli polisensor informasiya qabul qilinadi, qaysiki, bosh miya po’stlog’ining turli mintaqalariga yetib boradi. Ammo, oxirgi umumiy yo’ldan ayni vaqtda organizm uchun kerak bo’lgan eng asosiy va zaruriy axborot o’tadi (himoyalanish, ovqatlanish, seksual va hokazo). 3. Yonma-yon qavatlarda asab elementlarining miqdori teng emas. Masalan, ko’ruv tizimida reseptorlar soni 130 million, ko’zdan chiqadigan asabni tashkil qiluvchi neyronlarning soni esa faqat 1 mln 250 mingga yaqin yoki 100 marotaba kam. Bu nisbat miyaniing har qaysi yakka reseptorlarda vujudga kelgan impulsni qabul qila olmasligini ko’rsatadi. Buning biologik ahamiyati shundaki, informasiya markaziy asab tizimiga yetguncha bir necha marta saralanadi. Ko’p sonli qavatdan unsurlari kam qavatga o’tish jarayonida biologik nuqtai nazardan ikkinchi darajadagi informasiya olib qolinib, o’tkazilmaydi. Bu torayib boruvchi “voronka” (“Sherrington voronkasi”) miyaga o’tkaziladigan informasiyani kamaytiradi va muhim axborotlarnigina o’tkazadi. 4. Sensor tuzilmalarining vertikal va gorizontal bo’yicha farqlanishi. Bir bo’limda joylashgan analizator elementlarining funksional xossalari har xil bo’lishi mumkin. Masalan, ko’ruv analizatorining reseptor bo’limi rang sezuvchi kolbochkalardan va yorug’lik darajasini sezuvchi tayoqchalardan tashkil topgan. Bu farqlanish gorizontal farqlanish deyiladi.
Reseptorlar adekvat ta’sirlovchilarga, ya’ni evolyusiya davomida qaysi ta’sirotlarni sezishga maxsus moslashgan bo’lsa, o’sha ta’sirotlarga nisbatan juda yuksak darajada sezuvchanligi bilan farq qiladi. Qo’zg’alish ro’y berishi uchun zarur bo’lgan minimal miqdordagi energiyaga, ya’ni ta’sirot bo’sag’asiga qarab reseptorlarning qo’zg’aluvchanligi o’zagaradi. Reseptorlar noadekvat ta’sirlovchilar ta’sirida ham qo’zg’alishi mumkin. Masalan, ko’zga mexanik kuch (zarb) ta’sir etganda yorug’lik sezgisi kelib chiqadi (“ko’zim chaqnab ketdi” degan ibora shundan olingan), shu ta’sirlovchilar quloqqa ta’sir etganda tovush sezgisi paydo bo’ladi (“qulog’im shangillab ketdi”). Lekin noadekvat ta’sirlovchilar ta’sir etganda reseptorlar qo’zg’aluvchanligi ko’p marta kamroq bo’lib chiqadi. Reseptorlar qo’zg’aluvchanligi doimo bir xilda turmaydi. U bevosita reseptorlar funksional holati o’zgarganda, shuningdek markaziy asab tizimi (bosh miya po’stlog’i va to’rsimon formasiya) dan keluvchi impulslar ta’sirida ham o’zgarishi mumkin, bular reseptorlarning sezuvchanlik darajasini (“sozlanishini”) o’zgartiradi. I.M.Sechenov harakat analizatorini o’rganib, sezish reseptorlari bilan harakat reseptorlari bir-biriga bog’liqligini aniqlagan. Uning fikricha odamning sezgi a’zolari hayvonlar sezgi a’zolaridan tubdan farq qiladi. Keyinchalik I.P.Pavlov analizatorlarning morfofunksional tuzilishini o’rganib chiqdi. Uning fikricha ta’sirlovchilar ta’sirini qabul qilishda va asab impulslarini o’tkazishda qatnashadigan neyronlarning butun majmuasini, shuningdek bosh miya po’stlog’ining sensor (sezuvchi) neyronlarini yagona tizim hisoblab, ularni “analizator” deb nomlash mumkin. 2. Retseptorlarning tasnifi Reseptorlar ichki va tashqi reseptorlar deb ikkita katta guruhga bo’linadi. Ichki reseptorlar-interoreseptorlar-yuboradigan impulslar ichki a’zolarning holati haqida (visseroreseptorlar) va tana hamda undagi ayrim qismlarning fazodagi
vaziyati, harakati to’g’risida (vestibuloreseptorlar va proprioreseptorlar) xabar berib turadi. Tashqi reseptorlar- eksteroreseptorlar- tashqi olamdagi narsa va hodisalarning xossalari, ularning organizmga ta’siri to’g’risida signal berib turadi. Ba’zi reseptorlar organizmdan ancha olisdagi narsalardan keluvchi ta’sirotlarni qabul qila oladi. Bunday reseptorlar distant (masofadagi) reseptorlar deb ataladi. Ularga ko’ruv, eshituv, hid biluv reseptorlari kiradi. Boshqa- kontakt (teguvchi) reseptorlar faqat bevosita o’ziga tegib turgan, ya’ni reseptor tizimi yaqin yotgan narsalardan keluvchi ta’sirotlarni qabul qila oladi. Shunday qilib, reseptorlar-analizatorning periferik bo’limi hisoblanadi. Afferent (sezuvchi) neyronlar va o’tkazuvchi yo’llar analizatorning o’tkazuvchi bo’limini tashkil etadi. Bosh miya po’stlog’ining reseptorlardan qo’zg’alishni qabul qiladigan qismlari analizatorning markaziy oxirlari deb ataladi. Analizatorlarning o’tkazuvchi qismi-bu sensor (sezuvchi) asab tolasidan iborat bo’lib, u reseptordan ta’sirotni qabul qilib bosh miya po’stlog’ining proyeksion mintaqasiga yetkazadi. Analizatorning markaziy (po’stloqdagi) qismi bosh miya po’stlog’ining turli sohalarida joylashgan va sezuvchi asab markazlaridan iborat. Bu markazlarda muayyan sezgi a’zolaridagi reseptorlardan kelgan ta’sir tahlil qilinib, ularning mazmuniga ko’ra (sintez) javob reaksiyasi hosil bo’ladi (16.2.1-rasm). Analizatorning yuqorida aytilgan uchala qismining qaysi biri shikastlansa, muayyan sezgi a’zoning faoliyati buziladi. Reseptorlarning adekvat ta’sirlovchilariga qarab: 1) mexanoreseptorlar ta’sirlovchning mexanik energiyasini qabul qilishga moslashgan. Bunday reseptorlar teri, yurak-tomir tizimi, ichki a’zolar, tayanch harakat apparati, eshituv va muvozanatni saqlash tizimlariga xos. 2. Termoreseptorlar – harorat o’zgarishini sezadi. Issiqni va sovuqni sezuvchi termoreseptorlarning ko’p qismi terida joylashgan. Ichki a’zolar va markaziy asab tizimida ham shunday reseptorlar bor.
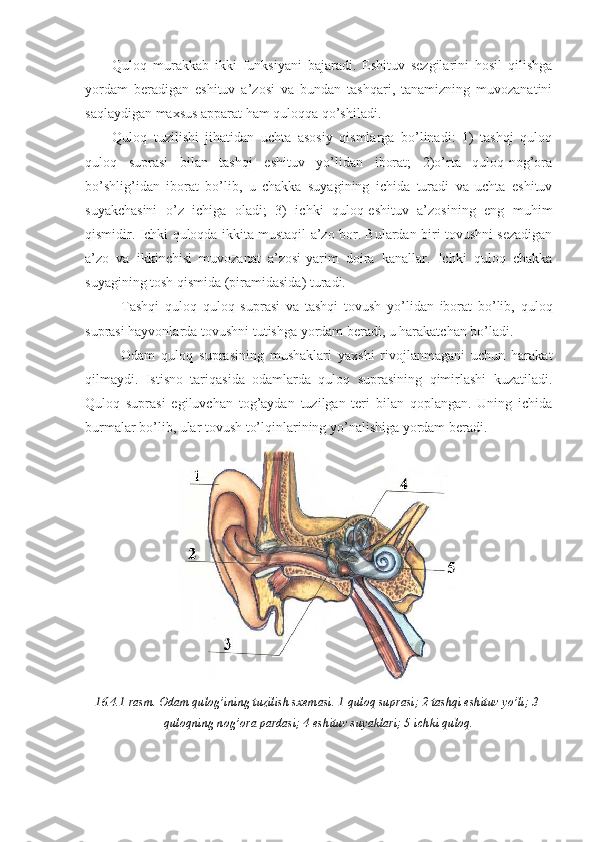
3. Xemoreseptorlar – kimyoviy omillar ta’siriga sezgir. Ular ta’m va hid sezuvchi sensor tizimlarning chet qismini tashkil qiladi. Bu tipdagi reseptorlar qon tomirlar tizimining turli qismlarida va ba’zi to’qimalarda ham uchraydi. 4. Fotoreseptorlar – nur energiyasini qabul qiladi. bu reseptorlar yorug’lik energiyasini ajratish va rang ko’rish imkonini beradi. 5. Og’riq (nosiseptiv) reseptorlari og’riqni paydo qiluvchi ta’sirotlarni qabul qiladi. bu sezgi organizmdagi reseptorlarning deyarli hammasiga o’ta ta’sir qilganda paydo bo’ladi. 16.2.1.-rasm. Bosh miya po’stlog’ida sezgi a’zolari markazlarining joylashuvi: 1-hid bilish markazi bosh miya po’stlog’i chakka qismining oldingi yuqori sohasida; 2-ko’ruv markazi ensa sohasida; 3-eshituv markazi bosh miya po’stlog’i chakka qismining pastki sohasida; 4-ta’m bilish markazi bosh miya po’stlog’i chekka qismining yuqori o’rta sohasida; 5-barmoqlar terisidagi paypaslash markazlari bosh miya po’stlog’i tepa qismining o’rta sohasida 6.Elektroreseptorlar – elektromagnit to’lqinlariga sezgir bo’lib, baliqlar va ba’zi amfibiylarning yon chizig’ida topilgan. Bularga elektrik energiyasini sezuvchi a’zolarining reseptorlari kiradi. 3. Sensor tizimlarning xususiy morfo-funksional tuzilishi